Knowledge on the high-ranking taxonomical groups of unicellular eukaryotes and their evolution; genetic, molecular and cellular criteria used in their classification; types of nutritional groups, acquisition of autotrophy and/or mixotrophy; reproduction; colonial unicellular organisms; metabolism of unicellular eukaryotes; responses to abiotic stress; biotechnological exploitation of heterotrophic, mixotrophic and autotrophic unicellular eukaryotes; unicellular eukaryotes used as biological models.
Comparison between classic and novel taxonomical classification systems involving genetic, molecular and ultrastructural characterization. Classification of unicellular eukaryotes proposed by Cavalier-Smith. Study of the chloroplast evolution and genes associated with its biogenesis as a starting point to understand the macroevolution of unicellular eukaryotes. Events of multiple endosymbiosis and their effect on the nutrition, metabolism and genomic organization. Adaptation and molecular and cellular responses to abiotic stress and extreme environments and use of this knowledge for the induction of metabolites with biotech-nological applications. Bioactive compounds. Use of knowledge of unicellular eukaryote biology for fighting parasitic diseases. Unicellular biological models for the study of the regulation of gene expression, signal transduction and metabolism of multicellular eukaryotic organisms.
Currently there is a growing consensus that the hierarchical taxonomy of Linnaeus is no longer adequate to describe evolutionary links due to the advancement of molecular taxonomy. This is particularly true for unicellular eukaryotes. As a rule, these cells have a rudimentary morphology, which has led to quite erroneous taxonomic classifications in the past, especially with regard to high ranking taxa . With the advent of molecular genetics, ultrastructural cell biology and bioinformatics, it became increasingly clear that the taxonomy of major groups of unicellular eukaryotes (erroneously referred to as "protists") had to be revised. Thus, in this subject, the latest developments are discussed, especially those concerning the tree of endosymbiotic events developed by Patrick Keeling, taking into account other alternatives such as the ones proposed by Thomas Cavalier-Smith.
Coordinator: João Varela (jvarela@ualg.pt)
Microalgae: Tamára Santos, Inês Maia, Marta Oliveira, Gabriel Bombo, Sofia Navalho, Bruno Pinto, Veronica Rossetto, Madalena Silva, Manuel Murtinho.
Bioactitivities: Marta Oliveira e Nicolle Prates
Subject matter is exposed in lectures in an interactive way, which are later complemented by theoretical-practical classes where the students discuss results of practical classes and how to write a scientific paper. In practical classes students learned methods for taxonomic classification of unicellular eukaryotes using molecular and microscopical methods. Evaluation is carried out via an exam (60% of the final grade), monograph (20%) and oral presentation of the monography (20%).
Introduction to the curricular unit. The 3 Domains of Life. The origin of eukaryotes. Comparison between eukaryotes and prokaryotes at biochemical level. The evolution of the taxonomy, which was initially based on morphological characters. The emergence of multidisciplinary taxonomy that involves the investigation and use of several criteria: morphological, physiological, biochemical, molecular and ultrastructural (cell biology). Review of the concepts of mono-, para- and polyphyly. Examples of convergent evolution. The dangers of using one criterion in taxonomy, namely molecular taxonomy. The problem of using rDNA for the definition of the tree of life. Ex. Microsporidia. How biochemistry and molecular biology helped define the 3 domains of life. Comparison between lipid stability with acyl and ether groups and its consequence in the survival at high temperatures.
Slides
Hagen (2012)
Delsuc et al. (2005)
Adl et al. (2005)
Cavalier-Smith et al. (2003)
Keeling (2009)
Keeling (2013)
How biochemistry and molecular biology helped define the 3 domains of life. Comparison between lipid stability with acyl and ether groups and its consequence in the survival at high temperatures (continuation of the last lecture).
The 6 major taxonomic groups of eukaryotes. Comparison with the 5 kingdoms of T. Cavalier-Smith. The 6 Kingdoms of Cavalier-Smith. The multilinear evolution proposed by Cavalier-Smith. The origin of life at 2.9-3.5 Gyears. The appearance of Negibacteria. The advantage of having 2 membranes in aquatic / aqueous environments. The appearance of the Posibacteria with the loss of the outer membrane. The appearance of the ancestor with neomuran and the derivation that gave rise to the Archaea.
Discussion and comparison of two papers, namely Hagen (2012) and Cavalier-Smith et al. (2004).
Discussion of themes and groups for the students' seminar. Discussion of basic terminology and concepts for the seminars.
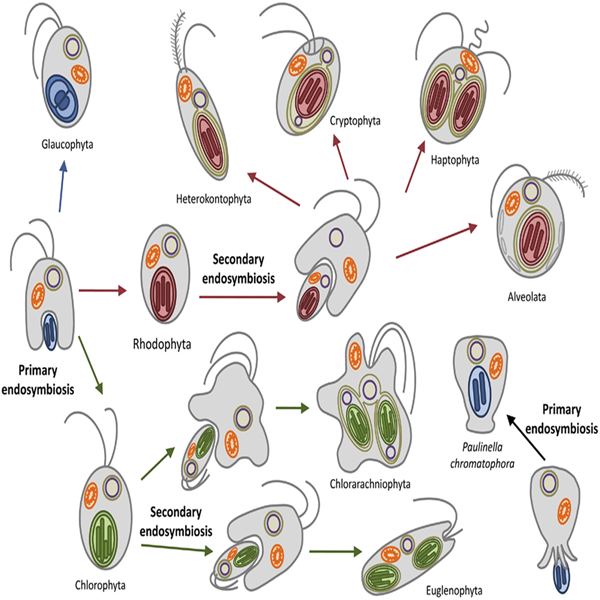
The evolution to an ancestral eukaryotic organism with endomembrane systems and the capture of a prokaryote that gave rise to mitochondria. The division of eukaryotes into two major evolutionary lines: unikonts and bikonts. Cellular, molecular and ultrastructural characteristics of unikonts. The lineage of the opisthokonts, which includes the Animalia, Fungi and the Choanozoa. Cellular, molecular and ultrastructural characteristics of bikonts. The Cabozoa, which includes the lineages Excavata (e.g., Euglenozoa) and Rhizaria (e.g., Foraminifera). Capture of cyanobacteria that eventually gave rise to the chloroplast by primary endosymbiosis and the appearance of the Archaeplastida / Plantae, which includes the Glaucophyta, Rhodophyta, Chlorophyta and Streptophyta. The evolution of this lineage that gave origin to the Rhodophyta with the production of pigments that allow the colonization of new habitats. Introduction to the megagroup Chromalveolata as proposed by Cavalier-Smith. "Kingdom" Chromista (e.g., Stramenopiles, Cryptophyta and Haptophyta) and Alveolata (e.g., dinoflagellates, cilliates and apicomplexans). Metagenomics and their need to study unicellular organisms not cultivable. Superkingdom Eukaryota, "Kingdom" Chromalveolata.
Slides
Keeling (2013)
Burki et al. (2016)
Brett et al. (1986)
Brett et al. (1994)
Archibald (2015)
Growth, isolation and morphological identification of microalgae via classic methods, such as plating and serial dilution.
Please go to Tutoria and log in to access its auxiliary file.
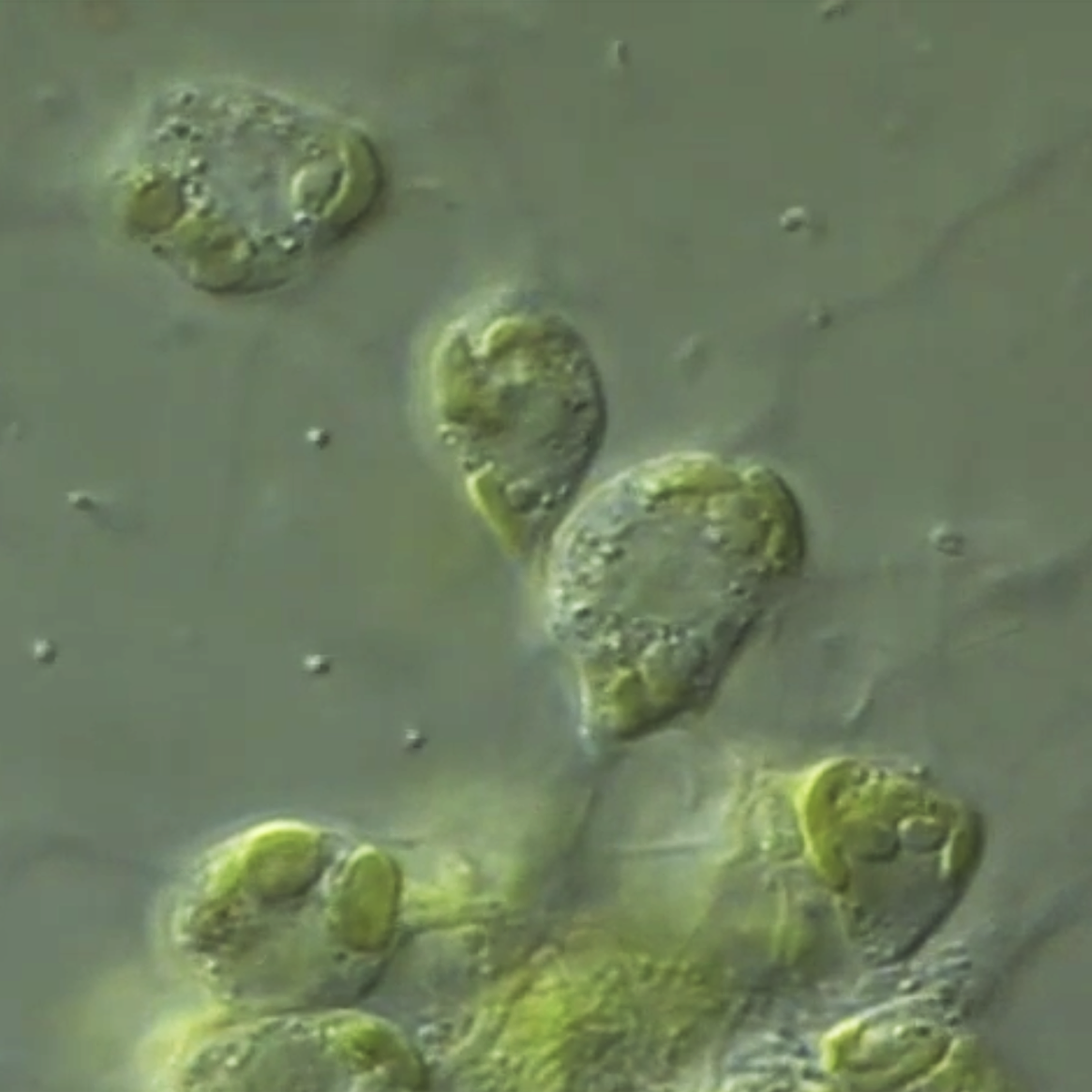
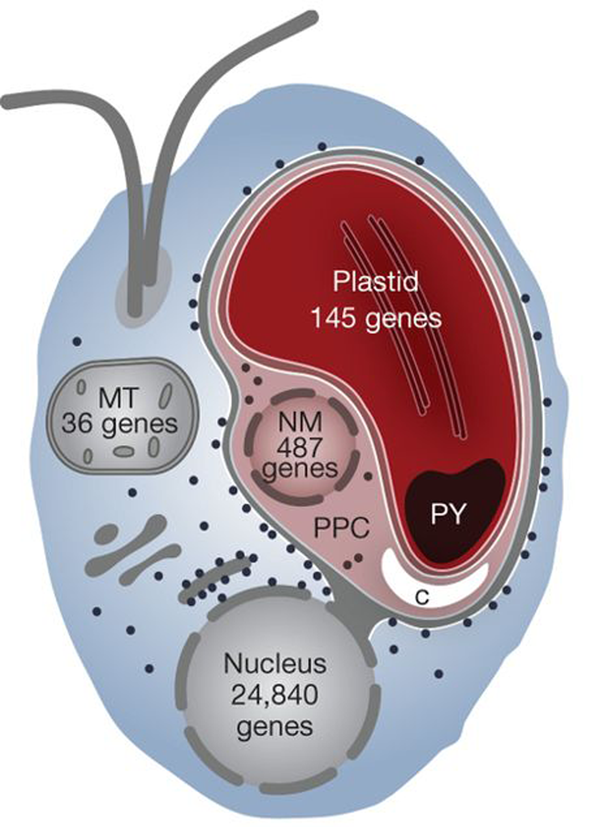
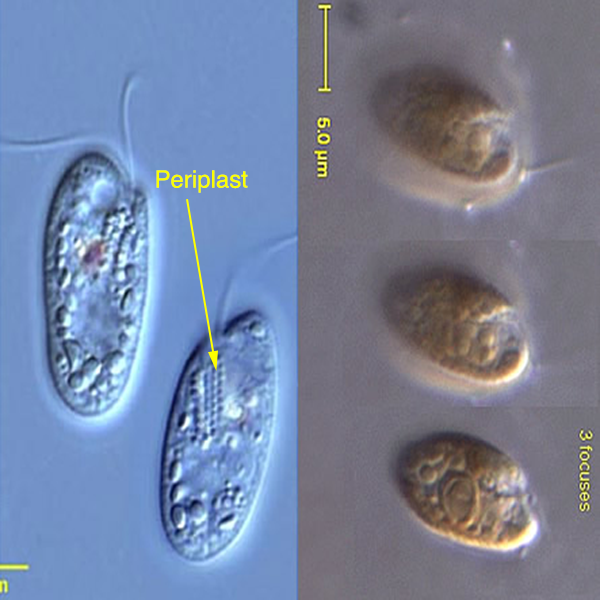
Superkingdom Eukaryota, "Kingdom" Chromalveolata. Doubts regarding the monophyly of this lineage. Characteristics common to the chromalveolates. The lineage Chromalveolata as an example of a taxonomic group with strong indications of HGT. The lineages Haptophyta and Cryptophyta as part of the lineage "Hacrobia", which current evidence does not support anymore as a monophyletic group. The lineage Rhizaria and Cercozoa as one of its most intriguing sub-lineages, which includes the Phylum Chlorarachniophyta. Cellular, biochemical characteristics and their likely evolutionary origin. Characteristics of the Haptophyta and Cryptophyta. Ultrastructural organization of the cryptophytes / cryptomonads. Chloroplast surrounded by 4 membranes. Definition of periplast and periplasm. The existence of ejectosomes and their probable function. Evolutionary origin of the 4 chloroplast membranes. The nucleomorph, the periplastidial space and the number of genomes present in cryptophytes. The nucleomorph of cryptophytes. The periplastidial space. Ultrastructural and biochemical evidence in favor of a secondary endosymbiosis (eukaryote-eukaryote).
Slides
Keeling (2013)
Brett et al. (1986)
Brett et al. (1994)
Riisberg et al. (2008)
Curtis et al. (2012)
Kamoun et al. (2005)
Discussion of questions and answers made by students about Hagen (2012) and Cavalier-Smith et al. (2004). Basic concept of a phylogenetic tree.
Evolutionary origin of the 4 chloroplast membranes of cryptophytes. The nucleomorph, the periplastidial space and the number of genomes present in cryptophytes. The periplastidial space and additional biochemical evidence in favor of a secondary endosymbiosis (eukaryote-rhodophyte). Comparison between Cryptophyta and Chlorarachniophyta. Stramenopiles / Heterokonta as a sub-lineage of the "Kingdom" Chromista as defended by Cavalier-Smith. Characteristics of heterokonts. Examples. Photoautotrophic heterokonts: Ochrophyta (previously also called Heterokontophyta). Eutigmatophytes, including Nannochloropsis.
Slides
Keeling (2013)
Brett et al. (1986)
Brett et al. (1994)
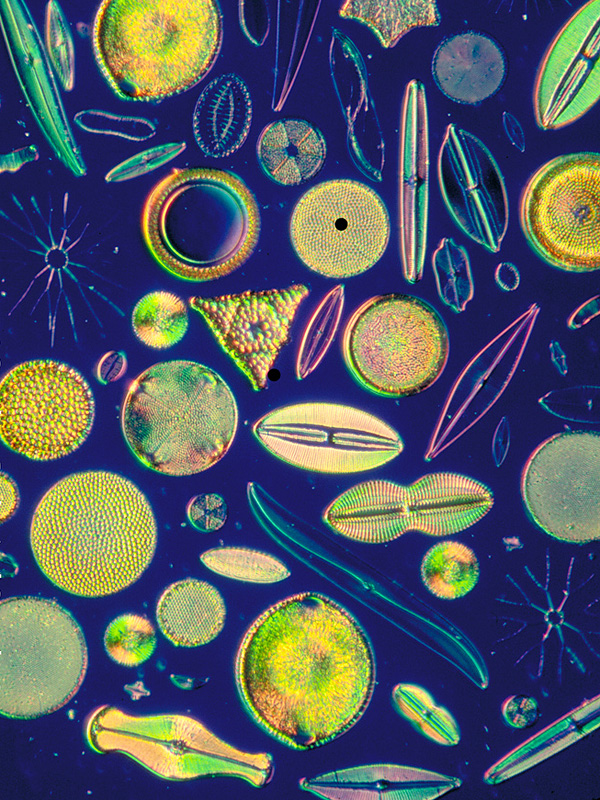
Superkingdom Eukaryota, "Kingdom" Chromalveolata.Photoautotrophic heterokonts: Ochrophyta (previously also called Heterokontophyta). Diatoms (Bacillariophyceae). The sub-lineages Pennatae and Centricae. The biodiversity of diatoms. Their impact on the trophic networks in the oceans and on the planet due to their contribution in the global fixation of CO2. Heterotrophic heterokonts: Pseudofungi. E.g., Phytophthora infestans. Characteristics of pseudofungi that are typical of true fungi. How biochemistry and molecular taxonomy led scientists to conclude that the pseudofungi are not fungi, but rather a heterotrophic lineage of heterokonts. Pseudofungi and fungi as an example of convergent evolution, which is reflected both morphologically and molecularly. Genes captured by Oomycetes that are fungal-like and their importance in their pathogenicity.
Slides
Keeling (2013)
Brett et al. (1986)
Brett et al. (1994)
Riisberg et al. (2008)
Curtis et al. (2012)
Kamoun et al. (2005)
Phylogeny. Determination of evolutionary distances with Poisson and Gamma corrections. Models of molecular evolution and phylogenetic inference methods.
The lineage Alveolata, with special emphasis on the Dinophyceae, although they also include the cilliates and apicomplexans. The "Chromalveolata hypothesis", which defines that only a single secondary endosymbiotic event gave rise to the "Chromista" (Stramenopiles + Hacrobia) and Alveolata as defended by Cavalier-Smith. Atypical cell cycle of dinoflagellates (zygotic meiosis) and comparison with the cell cycle of higher eukaryotes. Dinoflagellates as pathogens and toxic algae. How molluscs can protect themselves using toxins from dinoflagellates. The example of saxitoxin, which is considered to be a powerful chemical weapon that causes paralytic shellfish poisoning (PSP) symptoms. Lineage Archaeplastida: Glaucophyta, Rhodophyta, Chlorophyta.
Slides
Keeling (2013)
Curtis et al. (2012)
Kamoun et al. (2005)
Glasgow et al. (2001)
LaRonde et al. (1996)
Burki et al. (2012)
Leliaert et al. (2012)
Wisecaver & Hackett (2010)
Stoecker et al. (2009)
Ball et al. (2011)
Lineage Archaeplastida: Glaucophyta, Rhodophyta, Chlorophyta and Streptophyta (Charophyta + land plants). Glaucophyta and their chloroplasts: the cyanelles. Morphological diversity of the glaucophytes. The cyanelle ultrastructure, including the positioning of the phycobilisomes "outside" of the thylacoid (but in the most inner part of the chloroplast: the stroma). The phylum Chlorophyta and their evolutionary closeness to the Streptophyta, which includes the Charophyta and land plants (recapitulation). The prasinophytes as a "less" evolved group of marine chlorophytes. The freshwater "core chlorophytes" as the "more" evolved green algae. The Ulvophyceae (e.g., Ulva) as a multicellular group closely related to the "core chlorophytes". The Chlorodendrophyceae (e.g., Tetraselmis) as an intermediate group between the prasinophytes and the "core chlorophytes". The appearance of the eyespot / stigma as an early photoreceptor. Cell coverings of chlorophytes.
Slides
Keeling (2013)
Curtis et al. (2012)
Kamoun et al. (2005)
Glasgow et al. (2001)
LaRonde et al. (1996)
Burki et al. (2012)
Leliaert et al. (2012)
Wisecaver & Hackett (2010)
Stoecker et al. (2009)
Ball et al. (2011)
Continuation of classes about phylogenetic inference.
Classification of the green algae according to their morphology, ultrastructure (positioning of the basal bodies, connecting fibres and flagellar roots), type of cell coverings, mitosis and cytokinesis. The actual biochemical composition of microalgal cell walls (e.g., Chlorella). Comparison between Rhodophyta and Cryptophyta (cont.).
Slides
Keeling (2010)
Keeling (2013)
Leliaert et al. (2011)
Leliaert et al. (2012)
Gerken et al. (2013)
Domozych et al. (2012)
Lewis & McCourt (2004)
Molecular taxonomy of unicellular eukaryotes using phylogenetic inference.
Please go to Tutoria and log in to access its auxiliary files.
The Excavata lineage, with particular emphasis on Euglenophyta / Euglenozoa. Characteristics of euglenids (tri-membrane chloroplasts, the importance of the existence of chlorophyll b in chloroplasts). The importance of horizontal gene transfer for the understanding of the evolution of large groups of unicellular eukaryotic organisms, such as Excavata, Rhizaria, Chromalveolata and Archaeplastida. Endosymbiosis as a phenomenon that occurred several times throughout evolution, which resulted in the acquisition of photoautotrophy. Loss of autotrophy in groups that have become predators (e.g., cilliates) and / or parasites (e.g., Oomycetes, apicomplexans and Perkinsus). The photosynthetic megagroups as proposed by Keeling's team: Stramenopiles, Alveolata, Rhizaria, "Plants", Haptophyta and Cryptophyta. First evidence against the monophyly of Hacrobia. Mitochondria and chloroplasts display an evolutionary paradox concerning the frequency of endosymbiotic events. Archaeplastida / Plants are derived from a primary endosymbiotic event between an eukaryote and a prokaryote. Evidence in favour of this hypothesis. "Hacrobia" and euglenids as derived from a secondary endosymbiotic event. Photosynthetic rhizarians as derived from primary and secondary endosymbiotic events. Paulinella as the main example of the former case. Multiple secondary and tertiary endosymbiosis as drivers of the evolution within the lineage "Chromalveolata". Mizocytosis and kleptoplastidy. Conclusions on the acquisition of photosynthesis, HGT, old and new views on what a megagroup is, the importance of green algae as drivers of evolution in other lineages, and the distribution of storage polysaccharides as a way to determine which views on evolution seem to fit best the available biochemical data.
Slides
Keeling (2013)
Leliaert et al. (2011)
Leliaert et al. (2012)
Gerken et al. (2013)
Domozych et al. (2012)
Lewis & McCourt (2004)
Burki et al. (2008)
Burki et al. (2012)
Burki et al. (2016)
Nowack et al. (2014)
The monograph must contain the author's name, student's number, name of the course, master's degree, academic year, the name and address of the university and the place (Faro). Then it should contain a Summary, Introduction, Final Conclusion and Bibliography. It should also contain sections according to the theme to be developed between the introduction and final conclusion.
The Bibliographic Citation Rules in English or Portuguese must be faithfully followed. If you do not understand them, please talk to the coordinator of this curricular unit.
Deadline: 17 November, 2025 at 5 pm. Go to tutoria.ualg.pt, login, and look for Monograph inside the page of this curricular unit. Submit the monograph in PDF format and the presentation in PowerPoint format.
Dates of presentation of the monograph: 11th November 2025 at 16:00 (4 pm). Presentations should be no longer than 15 minutes.
Classification: average between the classification of the written monograph and classification of the oral presentation.